Đột biến knockout gen đích mã hóa “peroxisomal peptidase” gắn với tính kháng ngoài đồng bệnh hoại tử gây chết cây bắp
Mark Jung, Zhengyu Wen, Sabrina Humbert, et al. Targeted knockout of a host peroxisomal peptidase confers field resistance to maize lethal necrosis. PNAS April 30, 2026; 123 (18) e2535202123; https://doi.org/10.1073/pnas.2535202123

Bệnh hoại tử gây chết của cây bắp được viết tắt là MLN (Maize lethal necrosis), một bệnh siêu vi khá nghiêm trọng đe dọa an ninh lương thực ở Đông Phi. Tác giả tìm thấy một cơ chế chưa được biết trước đây, nhờ nó mà virus khai thác được một protein của bắp: peroxisomal peptidase để ngăn cản sự tự tái bản. Chính peptidase này hình thành nên một tổn thương di truyền rất báo động. Loại bỏ bằng cách sử dụng hệ thống chỉnh sửa hệ gen CRISPR-Cas mang lại khả năng kháng mạnh mẽ bệnh MLN. Dòng bắp Phi Châu chỉnh sửa gen vẫn duy trì tốt các tính trạng nông học cần thiết như bản gốc chưa chỉnh sửa, nhưng không có bệnh. Chiến lược đích đến như vậy cung cấp lộ trình bảo vệ năng suất cây bắp hiệu quả chống lại bệnh MLN đang đe gọa sản xuất, do vậy, bảo vệ được sinh kế của nông dân có quy mô sản xuất nhỏ, dễ bị tổn thương.
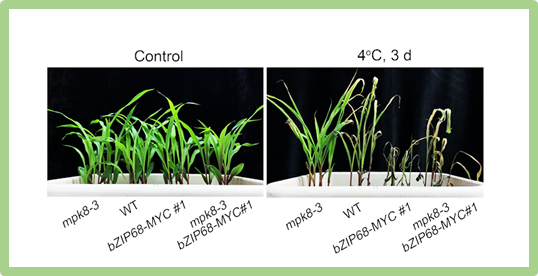
Bệnh hoại tử gây chết cây bắp (MLN) là bệnh nghiêm trọng do kết quả lây nhiễm kết hợp của virus MCMV (maize chlorotic mottle virus) và một potyvirus, có hầu hết trong bệnh khảm mía (SCMV). Bệnh này đe dọa nghiêm trọng đến an ninh lương thực vùng cận Saharan, châu Phi (SSA). Tác giả nghiên cứu một QTL chủ lực điều khiển tính kháng bệnh, định vị trên nhiễm sắc thể 6, có tên tiếng Anh là “maize lethal necrosis susceptibility locus 1” (ký hiệu qMLNS1), dẫn xuất từ giống bắp Thái Lan KS23-6. Kỹ thuật “Fine mapping” và kỹ thuật chỉnh sửa gen theo hệ thống “CRISPR-Cas9” những gen ứng cử viên trong quãng phân tử được thu hẹp còn 105 kb cho thấy peroxisomal peptidase là nguyên nhân gây nhiễm bệnh. Kết quả quan sát kính hiển vi confocal xác định vị trí của protein MLNS1 ở trong cơ quan peroxisomes. Tiến hành chỉnh sửa bằng kỹ thuật knockout đích đến của gen Mlns1 trong dòng bắp nhiễm bệnh CML536 từ điểm nóng châu Phi (SSA) xác định kết quả kháng so sánh với KS23-6 trên nghiệm thức thí nghiệm đồng ruộng tại Naivasha, Kenya. Dòng chỉnh sứa gen bằng knockout này khóa lại một cách chuyên nghiệp sự tích tụ siêu vi MCMV mà không ảnh hưởng gì đến SCMV. Dòng bắp chỉnh sửa gen biểu hiện không khác biệt năng suất hoặc không biểu hiện xấu đối với tính trạng nông học quan trọng trong điều kiện bệnh. Kết quả khẳng định cơ chế giữa enzyme trong cơ quan peroxisomal với sự lây nhiễm của virus. Tác giả còn xây dựng chiến lược nhanh, chỉnh sửa gen có khả năng mở rộng phục vụ kết hợp tính kháng MLN vào vật liệu di truyền, thu nhận một mô phỏng đấu tranh với những bệnh siêu vi khác ở loài cây trồng lương thực cơ bản của thế giới.
Xem https://www.pnas.org/doi/10.1073/pnas.2535202123

Hình: Kết quả “Fine mapping” những QTL đích điều khiển tính kháng bệnh MLN và xác định được gen đích trên nhiễm sắc thể 6.
GHI CHÚ
Quy trình chuẩn mực, đi từ Di truyền học số lượng (Quantitative Genetics) đến Sinh học phân tử và chức năng tế bào (Functional Genomics) để chứng minh chức năng của một gen đích điều khiển tính trạng (QTL chủ lực).
Dưới đây là phân tích hệ thống logic khoa học cho từng bước trong chuỗi nghiên cứu này và các hướng tiếp theo bạn cần triển khai để hoàn thiện bài báo/luận án:
1. Logic chuỗi liên hoàn trong nghiên cứu của bạn
Bước 1: Fine mapping (Bản đồ hóa độ phân giải cao)
Kết quả: Từ một vùng QTL rộng lớn (thường vài Mb), bạn đã thu hẹp được quãng phân tử xuống còn 105 kb.
Ý nghĩa: Khoảng cách 105 kb ở lúa hay các cây trồng khác thường chỉ chứa khoảng 10–15 gen ứng cử viên (candidate genes). Đây là tiền đề bắt buộc để bạn thực hiện bước tiếp theo là giải trình tự vùng này ở các dòng bố mẹ, phân tích tin sinh học để tìm ra gen MLNS1 (mang đột biến cấu trúc hoặc có sự khác biệt lớn về biểu hiện gen).
Bước 2: Định vị dưới tế bào bằng Kính hiển vi Confocal (Subcellular Localization)
Kết quả: Protein MLNS1 được xác định nằm trong peroxisomes (bào quan chuyên hóa tham gia vào quá trình oxy hóa acid béo, chu trình glyoxylate, và đặc biệt là giải độc các loài oxy hoạt tính - ROS hoặc tham gia tổng hợp phytohormone như jasmonic acid).
Ý nghĩa: Kết quả này cung cấp manh mối sinh hóa cực kỳ quan trọng. Nó gợi ý rằng QTL chủ lực này khả năng cao liên quan đến khả năng chống chịu stress (sốc nhiệt, hạn, mặn - nơi phát sinh nhiều ROS) hoặc quá trình chuyển hóa năng lượng/sinh trưởng.
Bước 3: Đánh giá chức năng bằng CRISPR-Cas9 Knockout
Kết quả: Tiến hành chỉnh sửa gen mã hóa protein MLNS1 để tạo dòng đột biến mất chức năng (knockout).
Ý nghĩa: Đây là bước tối cao (áp dụng định luật di truyền ngược) để khẳng định gen MLNS1 chính là gen chịu trách nhiệm cho kiểu hình của QTL chủ lực. Nếu dòng knockout (mlns1) bị mất hoặc giảm đáng kể tính trạng mong muốn so với dòng tự nhiên (Wild-type), hàm ý chức năng của gen đã được xác lập hoàn toàn.
2. Các thí nghiệm và phân tích cần làm tiếp theo để hoàn thiện nghiên cứu
Để dữ liệu đạt tiêu chuẩn công bố trên các tạp chí chuyên ngành quốc tế uy tín (Q1), bạn nên bổ sung các nội dung sau:
A. Phân tích phân tử dòng chỉnh sửa gen (CRISPR Lines Validation)
Giải trình tự vùng đích (Sanger Sequencing): Xác định chính xác kiểu đột biến ở thế hệ (ví dụ: mất 1 bp gây dịch khung, chèn 2 bp, hay mất đoạn lớn).
Sàng lọc dòng thuần không chứa T-DNA (Transgene-free): Ở thế hệ hoặc , sử dụng PCR với mồi đặc hiệu cho vùng promoter Cas9 hoặc gen kháng (như HPTII/Bar) để chọn ra các cây mang đột biến đồng hợp tử nhưng đã phân ly hoàn toàn khỏi cấu trúc vector CRISPR. Điều này đảm bảo kiểu hình quan sát được không bị ảnh hưởng bởi hiệu ứng ngoại lai.
B. Đánh giá kiểu hình sâu (Phenotyping) và Sinh hóa liên quan đến Peroxisome
Vì protein khu trú ở peroxisomes, bạn nên tập trung đo đạc các chỉ số sinh hóa đặc trưng của bào quan này trên cả 3 dòng: Dòng gốc (Wild-type), Dòng mang QTL (nếu có), và Dòng Knockout (mlns1):
Đo lường hàm lượng ROS và H₂O₂: Peroxisome là trung tâm xử lý . Hãy nhuộm DAB, NBT hoặc dùng các đầu dò huỳnh quang để xem dòng knockout có bị tích lũy độc chất ROS cao hơn khi gặp stress hay không.
Hoạt tính Enzyme chống oxy hóa: Khảo sát hoạt tính của các enzyme như Catalase (CAT), Superoxide Dismutase (SOD) và Peroxidase (POD).
Kiểm tra sự tích lũy acid béo: Nếu MLNS1 tham gia vào quá trình -oxy hóa acid béo ở peroxisome, hãy chạy GC-MS để phân tích thành phần lipid/acid béo ở hạt hoặc lá non.
C. Chứng minh bổ sung: Thí nghiệm cứu vãn kiểu hình (Complementation Test)
Để loại trừ hoàn toàn hiện tượng đột biến ngoài mục tiêu (off-target effect) của CRISPR, bạn nên làm thêm một bước Cứu vãn (Complementation):
Chuyển gen MLNS1 nguyên vẹn (Wild-type allele) dưới sự điều khiển của promoter bản địa của nó vào chính dòng đột biến knockout (mlns1).
Nếu dòng bổ sung gen này khôi phục lại được kiểu hình bình thường giống Wild-type, đây sẽ là bằng chứng tuyệt đối chứng minh không có sai sót off-target.
3. Gợi ý cấu trúc bài báo/báo cáo từ dữ liệu này
Map-based cloning: Thu hẹp vùng QTL xuống 105 kb và xác định MLNS1 là gen ứng cử viên sáng giá nhất dựa trên phân tích giải trình tự và mức độ biểu hiện (qPCR).
Functional characterization via CRISPR-Cas9: Tạo dòng knockout gen MLNS1 và chứng minh dòng này bị suy giảm kiểu hình nghiêm trọng so với dòng mang QTL trội/dòng gốc.
Mechanism insight: Định vị Confocal chứng minh protein hoạt động trong peroxisome, kết hợp dữ liệu sinh hóa (ROS/Catalase) để giải thích cơ chế tế bào tại sao gen này lại quyết định tính trạng của QTL chủ lực.